
Este estudio es el primer informe comunicando que el tamaño de la celdilla puede restringir la reproducción de Varroa destructor. Además nos hizo demostrar que un aspecto del entorno físico puede modular el comportamiento reproductivo de este parásito, dando lugar a nuevas hipótesis sobre las relaciones entre la nutrición y el éxito reproductivo de los ácaros.

Brood cell size of Apis Mellifera modifies the reproductive behavior of Varroa destructor
This study is the first report that cell width can restrict the reproduction of Varroa destructor. Moreover, we demonstrated that an aspect of the physical environment can modulate the reproductive behavior of this parasite, resulting in new hypotheses on relations between nutrition and reproductive success of the mites.
We undertook a field study to determine whether comb cell size affects the reproductive behavior of Varroa destructor under natural conditions. We examined the effect of brood cell width on the reproductive behavior of V. destructor in honey bee colonies, under natural conditions. Drone and worker brood combs were sampled from 11 colonies of Apis mellifera. A Pearson correlation test and a Tukey test were used to determine whether mite reproduction rate varied with brood cell width. Generalized additive model analysis showed that infestation rate increased positively and linearly with the width of worker and drone cells. The reproduction rate for viable mother mites was 0.96 viable female descendants per original invading female. No significant correlation was observed between brood cell width and number of offspring of V. destructor. Infertile mother mites were more frequent in narrower brood cells.
Introduction
Varroa destructor is an ectoparasitic mite of the Asian honey bee, Apis cerana that has become an important pest of the western honey bee (Apis mellifera) (De Jong et al. 1982b; Anderson and Trueman 2000). As this mite reproduces in honey bee brood cells, where temperature and humidity are tightly regulated by the bees, climate conditions appear unlikely to influence the life cycle of this parasite. However, Varroa reproduction can be influenced by host parameters (Ifantidis 1988; Eguaras et al. 1994).
Several studies have shown that the distribution of Varroa mites in brood combs differs significantly from a mathematical Poisson distribution and that it is aggregative (Donze´ et al. 1996; Floris 1991; Le Conte et al. 1990). The authors of these studies indicated that aggregation of Varroa female mites in brood cells suggests differences in the chemostimuli triggering infestation, such as higher production of kairomones from parasitized larvae, deposition of chemical trails, or emission of an aggregation pheromone by infesting mites. However, Fuchs (1988) suspected that fluctuations in the infestation rate are the reason for the non-Poisson distribution. This was later shown by Martin (1995), by controlling for variations in infestation level. When this influence was removed, he found that the female mites entered brood cells on a fairly random basis; the resulting clustering can be described by the Poisson distribution. Based on this result, Martin (1995) suggested that the over- dispersed distribution of mites found in other studies is likely to be a result of combining data sets with varying infestation levels.
Varroa mites reproduce and mate only in capped brood cells of bees, but only adult female mites live outside the brood cells (De Jong et al. 1982a). To infest cells and reproduce, Varroa mites walk off their adult-bee hosts onto the comb surface and then walk into cells where mature larvae are located (Boot et al. 1991). Various studies have investigated factors that could affect the infestation rate of V. destructor, such as local climate conditions (De Jong et al. 1984; Ritter and De Jong 1984), bee race (Correˆa- Marques et al. 2002; Moretto et al. 1991), infertility of adult female mites (Correˆa-Marques et al. 2003), absence of the queen (De Jong 1981) and the rate of removal of infested brood (Correˆa-Marques and De Jong 1998). In addition, chemical signals related to the invasion of brood cells have been reported. Le Conte et al. (1989) and Nazzi et al. (2004) identified putative attractants in extracts of bee larvae that affect the brood cell invasion behavior of V. destructor. Once mites contact late larval bee instars, they are apparently retained on the larval cuticle by hexane-extractable hydrocarbons (Rickli et al. 1994). It has also been demonstrated that physical signs affect brood cell invasion by Varroa mites (Kuenen and Calderone 2000). The distance from a bee larva to the rim of the brood cell appears to be a key factor in the process of cell invasion by Varroa (Boot et al. 1995; Goetz and Koeniger 1993). Also, old honey bee brood combs are more infested by Varroa mite than new brood combs (Piccirillo and De Jong 2004).
Cell size is an important determinant in the invasive behavior of V. destructor; more parasites per cell were found in worker brood cells that are individually raised above the surrounding cells (De Jong and Morse 1988; De Ruitjer and Calis 1988). Message and Gonc¸alves (1995) observed higher infestation and reproduction rates of V. destructor in European bee combs, when compared with Africanized combs, associated with their greater comb cell width. Piccirillo and De Jong (2003) found the same pattern among combs built by different subspecies of A. mellifera, when compared in the same colony. However, Taylor et al. (2008) reported that brood cell width had no significant effect on mite reproduction or infestation rate. These various studies of the effect of comb cell width were made by manipulating the brood nest, introducing combs with different cell sizes into colonies. However, how brood cell width affects the number of foundress mother mites per cell and its reproduction under natural conditions has not been studied previously.
The choice of a bee brood cell to begin reproduction is an important and critical moment in the life of the Varroa mite. In addition, the various reports of interaction between cell size and reproduction of the mite, in the interaction between V. destructor and it host A. mellifera; provide an excellent opportunity to study how selective pressures physical cues (in this case variability in cell width) can modify the reproductive behavior of an ectoparasite. Our objectives were to examine the natural variability in brood cell width in combs of A. mellifera and to analyze whether this factor affects the number of foundress mother mites per cell and its reproduction rate under natural conditions. The hypothesis tested was that brood cell width of A. mellifera is a key factor that affects the reproductive behavior of V. destructor under natural conditions. This hypothesis would be supported if infestation or reproduction rates vary with brood cell width.
Materials and methods
The study was conducted at the Laboratorio de Apicultura, J.J. Na´gera coastal station (3881000600S; 5783801000W) and at the Laboratorio de Artro´podos (Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata in Argentina. Drone cells were limited during the sampling period because we carried out the sampling during fall and spring 2007, when cold weather induces the bee colonies to restrict the brood area, especially the drone brood. Eleven colonies of A. mellifera maintained in Langstroth-type beehives were used for the experiments.
Population distribution of Varroa destructor
We examined the population distribution of V. destructor within brood cells of A. melli- fera. One sealed brood comb from each colony was intensively sampled. All of the brood cells of each comb were opened and the number of foundress mother mites (parasitic intensity) inside each cell was registered.
Morisita’s index (Brower and Zar 1977) was estimated according to the following equation:

where:
Is = Morisita’s dispersion index
N = Number of cells of A. mellifera inspected.
EX = Sum of the number of V. destructor foundress mother mites found within the inspected cells.
Based on the characteristics of this dispersion index, if:
Is = 1, the population distribution of V. destructor was adjusted to Poisson.
Is <1, the population distribution of V. destructor was normal.
Is >1, the population distribution of V. destructor was distributed according to negative binomial law.
To evaluate if the value of Is was significantly different from 1, the following equation
was employed:

If the value of F0 was greater than the tabulated F with n – 1 grades of freedom and 95% of confidence, it was concluded that Is was significantly different from 1. Infestation rate and mite distribution were analyzed for each colony. Infestation rate was estimated following De Jong (2005).
Brood cell width and its effect on the number of foundress mother mites invading cells and on its reproduction rate
Six colonies of A. mellifera were used. A drone brood comb was collected from one colony, while a worker brood comb (with no drone brood) was sampled from the remaining five colonies. Photographs of each comb were taken with a Kodak Easy Share C340 camera. The inner width of each sealed brood cell (distance between two parallel walls) was measured in each photograph using Image J software. Diameter distributions of worker and drone cells were graphed in frequency histograms.
All of the brood cells of each comb were opened and the number of foundress mother mites inside each cell was registered. To facilitate recording the data, a colored pin of different color was assigned to each opened cell according to the following criteria: green—zero mites; yellow—one mite; light blue—two mites; red—three or more mites. The combs were pho- tographed again with the pins. Using Image J software the two types of photographs (combs with sealed cells and those with opened cells with pins) were superimposed in order to relate the width of each sealed cell measured with the number of foundress mother mites and its reproduction rate. Reproduction rate of V. destructor was estimated only in worker brood cells. Only worker brood cells invaded by a single mother mite were analyzed, since multiple infestations can affect mite reproduction (Fuchs and Langenbachs 1989). Whenever possible, only brood that had been sealed for longer than 230 hours (at least at the yellow thorax stage of development) was collected. Mite eggs and protonymphs were not sexed, as this cannot be done reliably using external characteristics (Steiner 1988). Protonymphs and deutonymphs were distinguished by counting the number of sternal setae, according to Nannelli (1986). Reproduction rate was calculated for foundress mother mites according to Ifantidis (1984), by dividing the total progeny of the mites by the number of mites of the parental generation.
To determine whether brood cell width affected the number of V. destructor foundress mother mites in the brood, a generalized additive model (GAM) (Hastie and Tibshirani 1990; Zuur et al. 2007), which is based on Binomial and Poisson distributions, was used for both worker and drone cells.
To determine whether brood cell size affected V. destructor reproduction rate, we used a Pearson’s correlation and a Tukey test to calculate the relationship between the total number of mites infesting each brood cell (original adult and offspring) and the width of the brood cell. Each foundress mother mite was classified into one of the following three categories: (1) viable mother mite (a mother mite that produced live female and male offspring); (2) inviable mother mite progeny (a mother mite that produced incomplete offspring due to dead female or male offspring or only a male produced); (3) infertile mother mite (mother mite did not reproduce). Tukey test was used to compare cell width means whenever a one-way ANOVA indicated significant differences between cell width means for the three categories of mother mites.
Results
Population distribution of Varroa destructor
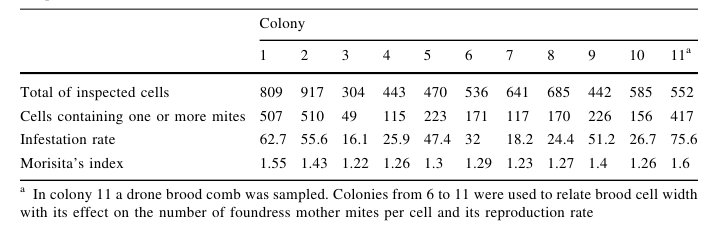
From the eleven colonies, 6,394 worker brood cells were inspected. The distribution of V. destructor adjusted to the negative binomial distribution in all colonies. Morisita’s index was significantly greater than one (P \0.05), in all colonies (Table 1). When data for all colonies were pooled, Morisita’s index was 1.344 and was significantly different from 1 (F0 = 1.32 [Ft; P \0.001). Infestation rates for each colony varied from 16.1 to 75.6% (Table 1). The mean infestation rate (±standard deviation) for all colonies was 39.6 ± 19.8% Frequency distribution of mean numbers of mites per cell per colony and standard deviations for pooled data are shown in Fig. 1.
Brood cell width and its effect on the number of foundress mother mites invading cells and on its reproduction rate
A total of 2,899 worker cells and 552 drone cells from the six colonies were examined. The mean width of the worker brood cells was 5.26 ± 0.51 mm (range = 4.17–6.86 mm), while for the drone brood cells it was 6.4 ± 0.5 mm (range = 5.05–8.07 mm). Worker and drone cell widths overlapped at from 5.5 to 6.8 mm (Figs. 2 and 3). A unimodal distribution of cell width was observed for both castes, with low relative frequencies for the extreme values (Figs. 2 and 3). Drone cell mean widths were 1.22 times greater than worker cell mean widths, although some worker cells were larger than the mean drone cell size.
Parasite intensities were 1.46 ± 0.34 foundress mother mites/worker cell (mean infestation frequency = 40.87%) and 2.16 ± 0.56 foundress mother mites/drone cell (mean infestation frequency = 75.36%).
GAM analysis for worker brood cells was significant (v2 = 104.4; df = 2,786; P \2e-16); it explained 5.15% of the deviance. The results presented here showed that the probability to find parasites inside cells worker brood cells was positively correlated with worker brood cell width. The relation was sigmoidal: the probability to find parasites inside worker brood cells increased lineally from cells of 4 mm to 5.9 mm where it became asymptotic (Fig. 2). GAM analysis for the drone brood cells was significant (v2 = 163.4; df = 2,399; P \2e-16); it explained 14.3% of the deviance. The number of parasites inside drone cells was positively correlated with cell width. The relation was sigmoidal: the number of foundress mother mites of V. destructor increased linearly from 5 to 6.8 mm, after which it became asymptotic (Fig. 3).
Table 1 Number of inspected cells, parasitized cells, infestation rate and Morisita’s index for each colony sampled


Foundress mother mites per cell varied with the proportion of available worker and drone brood cells (Figs. 2 and 3). The number of mites per cell increased positively and linearly with the width of worker and drone cells, until it became constant due to a decrease in the availability of wider cells.
Ninety-five of the 2,899 worker brood cells that we examined contained a single mother mite (3.3% single infestation). No significant correlation (r = 0.25; P = 0.2) was observed between cell width and the number of V. destructor offspring. Fifty-eight percent of the

singly-infested worker cells had viable offspring, 26% had inviable offspring and 16% contained infertile mother mites. The rate of increase for viable mother mites was 0.96 viable female descendants. Mean cell width was significantly different among the cate- gories of mother mite progeny (ANOVA, F = 6.343, P = 0.003); the mean widths of cells with infertile females, and viable and inviable progeny were 4.92, 5.4 and 5.38 mm, respectively.
Discussion
According to Poulin (1998), parasite habitats are not spatially continuous; they consist of discrete cells or ‘‘islands’’. Hosts constitute patches of suitable habitat in an otherwise inhospitable environment. In addition, parasites are aggregated among available hosts, such that most hosts harbor no or few parasites and few hosts harbor many parasites. The tendency towards parasite aggregation suggests that similar processes may be acting to generate the same pattern in different host-parasite systems, i.e., reproduction of parasites within hosts, density-dependent parasite mortality and parasite-induced host mortality (Anderson and Gordon 1982).
We found the population distribution of V. destructor within brood combs of A. mellifera to be aggregated; it was explained partially by the distribution of cell widths. Other studies have shown that the distribution of Varroa mites in cells differs significantly from a random distribution and that it is aggregative (Donze´ et al. 1996; Floris 1991; Le Conte et al. 1990). These authors indicated that the aggregation of Varroa female mites in brood cells suggests differences in the chemostimuli triggering infestation, such as higher production of kairomones by parasitized larvae, deposition of chemical trails, and pro- duction of an aggregation pheromone by infesting mites. However, brood cell dimensions also play an important role in the cell invasion of V. destructor (De Jong and Morse 1988; De Ruitjer and Calis 1988; Message and Gonc¸alves 1995; Piccirillo and De Jong 2003). We found that the distribution of cell widths among worker and drone cells in a brood comb of A. mellifera under natural conditions is not uniform and that it is highly variable, resulting in habitat patches that can be preferentially colonized by V. destructor. We found the variability in the width of cells to have a direct effect on the aggregation of V. destructor within brood combs. Although Martin (1995) concluded that the overdi- spersed distribution of mites is likely to be a result of combining data sets with varying infestation rates, we found that all our colonies had overdispersed mite distributions, independent of the infestation rates. Moreover, it was only used data from one brood for each colony, so it was impossible that the overdispersed distribution found in this study was the result of combining data sets. There is controversy about which factors, including physical and chemical elements, are important in triggering the invasion of brood cells by V. destructor and consequently affect its aggregation. Calderone and Lin (2001) proposed four V. destructor invasion models based on physical interactions, chemical interactions, both physical and chemical interaction, or due to passive mechanisms (Message and Gonc¸alves 1995). Kuenen and Calderone (2000) proposed that physical factors are the most important. Among physical factors, brood cell size has been well studied; it has been documented that this factor can affect the invasion and reproduction rates of Varroa mites. Higher parasite infestations were found to occur in worker cells that were individually raised above the surrounding brood cells (De Jong and Morse 1988; De Ruitjer and Calis 1988). Piccirillo and De Jong (2003) found that when comparisons were made within the same colony, the largest category of brood cells, which were Carniolan bee combs (mean width of 5.3 mm), were about 38% more infested than Italian comb brood cells (mean width 5.15 mm), which in turn were about 13% more infested than the self-built Afri- canized combs (mean width of 4.8 mm). Message and Gonc¸alves (1995) found similar results when infestation rates of V. destructor were compared between European and Africanized honey bee combs in colonies of Africanized bees. Worker brood that develops in large cells should receive more visits from nurse workers, increasing the probability of the mites transferring to the cells to infest the brood. These factors could increase the probabilities of the female mites entering into the cells, and in turn such cells would contain a larger bee larva and consequently offer more food to the mite.
We found that brood cell width in A. mellifera colonies affected the invasion and reproduction rate of V. destructor under natural conditions. The parasite invasion rate increased positively and linearly with the width of worker and drone brood cells (probably because brood developed in large cells will receive more visits from nurse workers, increasing the probability of the mites transferring to the cells to infest the brood), until it became constant when it reached a certain cell width. Moreover, it was found that infertile mother mites were more common in the smaller cells. Earlier studies had reported that larger cells promoted higher parasite reproductive success than smaller ones (Fuchs and Langenbach 1989; Message and Gonc¸alves 1995; Schulz 1984), but there are no reports that cell width can stop Varroa mite reproduction. Taylor et al. (2008) found that brood cell width had no significant effect on mite reproduction and they concluded that factors that are attributable to the race of bee, in combination with small brood cells, reduce the mite infestation and production of offspring. We found no correlation between cell width and the number of offspring of V. destructor. Nevertheless, cells with infertile mother mites (without offspring) were significantly smaller than cells with viable and unviable offspring. These results showed that cell width can drastically influence the invasion and the reproduction rates of the mites. Donze´ and Guerin (1994) reported that V. destructor had several behavioral adaptations to ensure its reproduction inside brood cells, because the free space inside the cells is continually changing during the bee’s metamorphosis. In addition, Martin and Kryger (2002) showed that a reduced space between the bee pupa and the cell wall in worker brood cells negatively impacts on the reproductive success of the mite because it can not move normally inside cells. Cell width plus a stress effect (e.g., nutrition effect) could be the connection between the lack of mite reproduction and the size of the cell that we found. Previous researches have reported that the nutrition quality improves mite reproduction (Rodriguez et al., 1961; Steiner et al., 1995). It was reported that Varroa mite reproduction can be suppressed under stress conditions due to severe reproductive competition, which can result in egg resorption by the foundress mother mite (Steiner et al., 1995). Also, Alberti and Hanel (1986) showed that mites that had not laid eggs often had problems with prosperm maturation in the reproductive tract of the female mites. Like egg resorption, problems with prosperm maturation could be due to stress conditions. Tewarson (1983) reported that reproduction of this parasite occurs only after intake of a number of blood meals; some bee hemolymph proteins are absorbed without digestion. Steiner et al. (1995) demonstrated that oogenesis is faster in drone brood cells than in worker brood cells, probably due to greater food availability for the foundress mother mite. Based on this line of reasoning, the smaller cells could contain smaller larvae, which would provide a smaller quantity of nutritients for the initiation of mite reproduc- tion. Harris and Harbo (1999) suggested that mites that had not laid eggs had problems with mating or a failure of the prosperm to mature in the reproductive tract of the female mites. However, they were not able to distinguish between nonmating and failure of sperm maturation.
If the cell width distributions found in this study (where bigger cells are less numerous) are considered, there could be a trade-off between the time necessary to search for optimal cells and the higher reproductive success obtained in such cells: Varroa females try to find bigger cells because they do not reproduce as efficiently in the small ones (probably because of a food stress factor); although finding big brood cells would insure better conditions for parasite reproduction, the associated costs with searching for infrequent cells would not be compensated by the benefits of finding such cells, making this strategy inefficient (Optimal Foraging, Charnov 1976). This hypothesis is supported by the evi- dence presented by Boot et al. (1994), who found that the invasion rate increases with the number of brood cells available. Moreover, to search for bigger cells, with the higher number of foundress mother mites inside, as we report here, could result in a reduction in the reproductive success of Varroa female mites because of a negative density-dependent effect (Eguaras et al. 1994; Fuchs and Langenbach 1989). These hypotheses could be tested in future experiments.
![]() This study is the first report that cell width can restrict the reproduction of V. destructor. Moreover, we demonstrated that an aspect of the physical environment can modulate the reproductive behavior of this parasite, resulting in new hypotheses on relations between nutrition and reproductive success of the mites.
This study is the first report that cell width can restrict the reproduction of V. destructor. Moreover, we demonstrated that an aspect of the physical environment can modulate the reproductive behavior of this parasite, resulting in new hypotheses on relations between nutrition and reproductive success of the mites.
![]() Diese Studie ist der erste Bericht dass die Zellgröße die Vermehrung von V. destructor beschränken kann. Darüber hinaus konnten wir zeigen, dass ein Aspekt der physischen Umgebung die Fortpflanzungsverhalten dieser Parasiten modulieren können, was zu neuen Hypothesen führt über die Beziehungen zwischen Ernährung und Fortpflanzungserfolg der Milben.
Diese Studie ist der erste Bericht dass die Zellgröße die Vermehrung von V. destructor beschränken kann. Darüber hinaus konnten wir zeigen, dass ein Aspekt der physischen Umgebung die Fortpflanzungsverhalten dieser Parasiten modulieren können, was zu neuen Hypothesen führt über die Beziehungen zwischen Ernährung und Fortpflanzungserfolg der Milben.
![]() Este estudio es el primer informe comunicando que el tamaño de la celdilla puede restringir la reproducción de Varroa destructor. Más encima nos hizo demostrar que un aspecto del entorno físico puede modular el comportamiento reproductivo de este parásito, dando lugar a nuevas hipótesis sobre las relaciones entre la nutrición y el éxito reproductivo de los ácaros.
Este estudio es el primer informe comunicando que el tamaño de la celdilla puede restringir la reproducción de Varroa destructor. Más encima nos hizo demostrar que un aspecto del entorno físico puede modular el comportamiento reproductivo de este parásito, dando lugar a nuevas hipótesis sobre las relaciones entre la nutrición y el éxito reproductivo de los ácaros.
Acknowledgments The authors thank the UNMDP and CONICET for financial support. This research was supported by an ANPCyT, Pict 07 grant to M. E. We thank Dr. Norma Sardella for her criticisms and suggestions and to Dr. Elena Ieno for her help in the statistical analysis.
References
Alberti G, Hanel H (1986) Fine structure of the genital system in the bee parasite, Varroa jacobsoni (Gamasida: Dermanyssina) with remarks on spermiogenesis, spermatozoa and capacitation. Exp Appl Acarol 2:63–104
Anderson R, Gordon D (1982) Processes influencing the distribution of parasite numbers within host populations with special emphasis on parasite-induced host mortalities. Parasitology 85:373–398 Anderson D, Trueman J (2000) Varroa jacobsoni is more than one species. Exp Appl Acarol 24:165–189
Boot W, Calis J, Beetsma J (1991) Invasion of Varroa mites into honeybee brood cells; when do brood cells attract Varroa mites? Proc Exp Appl Entomol 2:154–156
Boot W, Sisselaar D, Calis J, Beetsma J (1994) Factors affecting invasion of Varroa mites into honey bee brood cells. Bull Entomol Res 84:3–10
Boot W, Driessen R, Calis J, Beetsma J (1995) Further observations on the correlation between attrac- tiveness of honey bee brood cells to Varroa jacobsoni and the distance from larva to cell rim. Entomol Exp Appl 76:223–232
Brower J, Zar J (1977) Field and laboratory methods for general ecology. Wm. C. Brown Company Publishers, Iowa
Calderone NW, Lin S (2001) Behavioural responses of Varroa destructor (Acari: Varroidae) to extracts of larvae, cocoons and brood food of worker and drone honey bees, Apis mellifera (Hymenoptera: Apidae). Physiol Entomol 26:341–350
Charnov EL (1976) Optimal foraging: the marginal value theorem. Theor Popul Biol 9:129–136Correˆa-Marques MH, De Jong D (1998) Uncapping of worker bee brood, a component of the hygienic
behavior of Africanized honey bees against the mite Varroa jacobsoni Oudemans. Apidologie 29:283– 289
Correˆa-Marques MH, De Jong D, Rosenkranz P, Gonc¸alves LS (2002) Varroa-tolerant Italian honey bees introduced from Brazil were not more efficient in defending themselves against the mite Varroa destructor than Carniolan bees in Germany. Genet Mol Res 1:153–158
Correˆa-Marques MH, Medina LM, Martin SJ, De Jong D (2003) Comparing data on the reproduction of Varroa destructor. Genet Mol Res 2:1–6
De Jong D (1981) Effect of queen cell construction on the rate of invasion of honeybee brood cells by Varroa jacobsoni. J Apic Res 20:254–257
De Jong D (2005) Workshop sobre Control de la Varroosis en Climas Subtropicales. 27 y 28 de junio, Salta Argentina
De Jong D, Morse RA (1988) Utilization of raised brood cells of the honey bee, Apis mellifera (Hyme- noptera: Apidae), by the mite Varroa jacobsoni (Acarina: Varroidae). Entomol Gen 14:103–106
De Jong D, De Jong PH, Gonc¸alves LS (1982a) Weight loss and other damage to developing worker honeybees from infestation with Varroa jacobsoni. J Apic Res 21:165–167
De Jong D, Morse RA, Eickwort GE (1982b) Mites pests of honey bees. Annu Rev Entomol 27:229–252 De Jong D, Gonc¸alves LS, Morse RA (1984) Dependence on climate of the virulence of Varroa jacobsoni.
Bee World 65:117–121
De Ruijter A, Calis J (1988) Distribution of Varroa jacobsoni female mites in honey bee worker brood cells of normal and manipulated depth (Acarina: Varroidae). Entomol Gen 14:107–109
Donze´ G, Guerin MP (1994) Behavioral attributes and parental care of Varroa mites parasitizing honeybee brood. Behav Ecol Sociobiol 34:305–319
Donze´ G, Hermann M, Bachofen B, Guerin M (1996) Effect of mating frequency and brood cell infestation rate on the reproductive success of the honeybee parasite Varroa jacobsoni. Ecol Entomol 21:17–26 Eguaras M, Marcangeli J, Fernandez N (1994) Influence of the parasitic intensity on Varroa jacobsoni Oud.
reproduction. J Apic Res 33:155–159
Floris I (1991) Dispersion indices and sampling plans for the honeybee (Apis mellifera ligustica Spin.) mite Varroa jacobsoni Oud. Apicoltura 7:161–170
Fuchs S (1988) The distribution of Varroa jacobsoni on honeybee brood combs and within brood cells as a consequence of fluctuation rates. In: Cavalloro R (ed) European research on varroatosis control: proceedings of a meeting of the EC expert0s group, Bad Homburg, 15–17 October 1986. Balkema, Rotterdam, pp 73–76
Fuchs S, Langenbach K (1989) Multiple infestation of Apis mellifera L. brood cells and reproduction of Varroa jacobsoni Oud. Apidologie 20:117–130
Goetz B, Koeniger N (1993) The distance between larva and cell opening triggers brood cell invasion by Varroa jacobsoni. Apidologie 24:67–72
Harris J, Harbo J (1999) Low sperm counts and reduced fecundity of mites in colonies of honey bees (Hymenoptera: Apidae) resistant to Varroa jacobsoni (Mesostigmata: Varroidae). J Econ Entomol 92(1):83–90
Hastie T, Tibshirani R (1990) Generalized additive models. Chapman and Hall, London
Ifantidis MD (1984) Parametres of the population dynamics of the Varroa mite on honeybees. J Apic Res 23:227–233
Ifantidis MD (1988) Some aspects of the process of Varroa jacobsoni mite entrance into honey bee (Apis mellifera) brood cells. Apidologie 19:387–396
Kuenen L, Calderone N (2000) Varroa mite infestation in elevated honey bee brood cells: effect of context and caste. J Insect Behav 13(2):201–213
Le Conte Y, Arnold G, Trouiller J, Masson C, Chappe B, Ourisson G (1989) Attraction of the parasitic mite Varroa to the drone larvae of honey bees by simple aliphatic esters. Science 245:638–639
Le Conte Y, Arnold G, Desenfant P (1990) Influence of the brood temperature and hygrometry variations on the development of the honey bee ectoparasite Varroa jacobsoni. Environ Entomol 19:1780–1785
Martin S (1995) Ontogenesis of the mite Varroa jacobsoni Oud. In drone brood of the honeybee Apis mellifera L. under natural conditions. Exp Appl Acarol 19:199–210
Martin S, Kryger P (2002) Reproduction of Varroa destructor in South African honey bees: does cell space influence Varroa male survivorship? Apidologie 33:51–61
Message D, Gonc¸alves LS (1995) Effect of the size of worker brood cells of africanized honey bees on infestation and reproduction of the ectoparasitic mite Varroa jacobsoni Oud. Apidologie 26:381–386 Moretto G, Gonc¸alves LS, De Jong D (1991) The effects of climate and bee race on Varroa jacobsoni Oud.
infestations in Brazil. Apidologie 22:197–203
Nannelli R (1986) Caratteri morfologici essenzialli per una rapida identificazione dei diversi stadi di Varroa jacobsoni Oud. Apicoltura 2:95–119
Nazzi F, Milani N, Della Vedova G (2004) A semiochemical from larval food influences the entrance of Varroa destructor into brood cells. Apidologie 35:403–410
Piccirillo G, De Jong D (2003) The influence of brood comb cell size on the reproductive behavior of the ectoparasitic mite Varroa destructor in Africanized honey bee colonies. Genet Mol Res 2:36–42 Piccirillo G, De Jong D (2004) Old honey bee brood combs are more infested by the mite Varroa destructor
than are new brood combs. Apidologie 35:359–364
Poulin R (1998) Evolutionary ecology of parasites: from individuals to communities. Chapman and Hall, London
Rickli M, Diehl PA, Guerin PM (1994) Cuticle alkanes of honeybee larvae mediate arrestment of bee parasite Varroa jacobsoni. J Chem Ecol 20:2437–2453
Ritter W, De Jong D (1984) Reproduction of Varroa jacobsoni Oud. in Europe, the middle East and tropical South America. Z Angew Entomol 98:55–57
Rodriguez J, Wade G, Claude F (1961) The nutrition of Macrocheles muscaedomesticae (Acarina: Mac- rochelidae) in relation to its predatory action on the house fly egg. Ann Entomol Soc Am 54(6):782– 788
Schulz A (1984) Reproduktion und Populationsentwicklung der parasitischen Milbe Varroa jacobsoni Oud. in Abhfingigkeit vom Brutzyklus ihres Wirts Apis mellifera L. Apidologie 15:401–420
Steiner J (1988) Sex discrimination based on external structures in nymphal and adult Varroa jacobsoni mites (Acarina: Varroidae). Entomol Gen 14:133–138
Steiner J, Diehl PA, Vlimant M (1995) Vitellogenesis in Varroa jacobsoni, a parasite of honey bees. Exp Appl Acarol 19:411–422
Taylor M, Goodwin R, McBrydie H, Cox H (2008) The effect of honey bee worker brood cell size on Varroa destructor infestation and reproduction. J Apic Res/Bee World 47(4):243–246
Tewarson NC (1983) Nutrition and reproduction in the ectoparasitic honey bee (Apis sp.) mite, Varroa jacobsoni. Thesis, Eberhard-Karls-University, Tu¨bingen, Germany
Zuur A, Ieno E, Smith G (2007) Analysing ecological data. Springer, New York